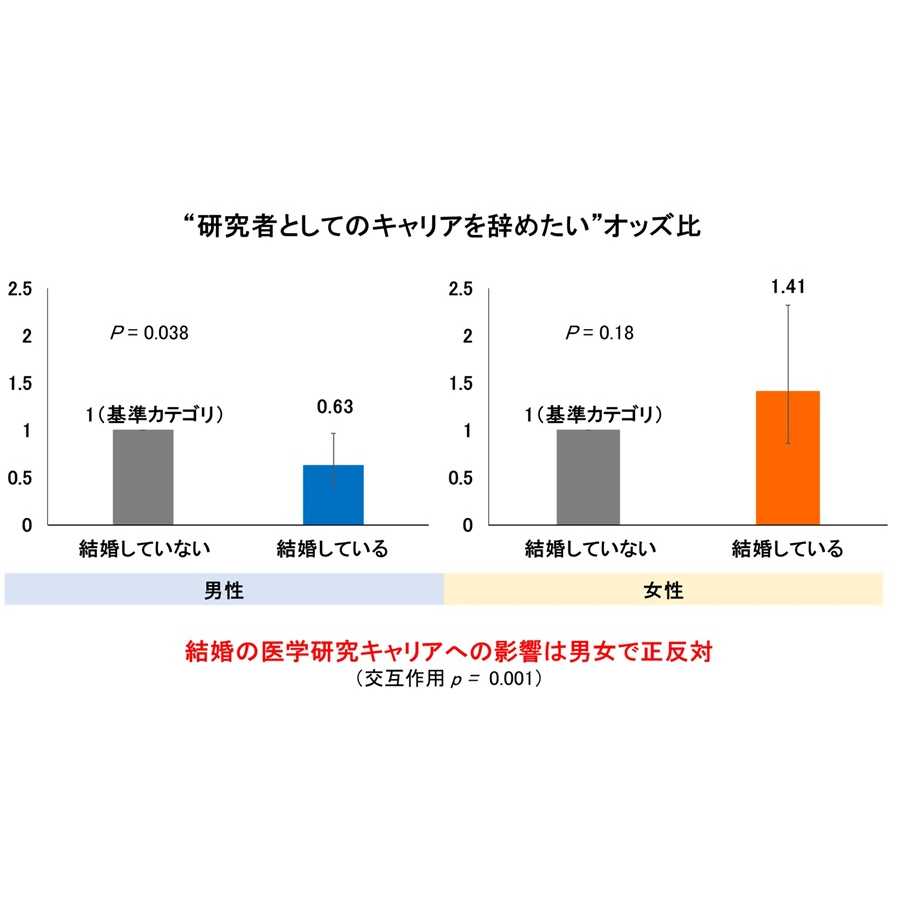
研究内容
本研究では、ヒト発生におけるUTYの役割を明らかにするため、Y染色体を持つヒト胚性幹細胞(ES細胞)
*5を用いて解析を行いました。ヒト胚性幹細胞は、初期胚に由来し、未分化な状態を保ったままさまざまな細胞へと分化できる性質(多能性)を持つ細胞です。
Y染色体は繰り返し配列が多く、遺伝子の発現量も低いことから、その機能解析はこれまで困難とされてきました。本研究では、ゲノム編集技術
*6によりUTYに人工タグ
*7を付加し、細胞内での局在やゲノム上での結合領域を高精度に解析しました。
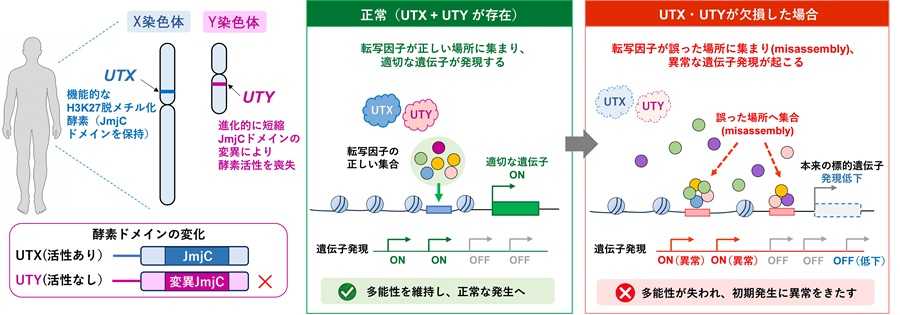
その結果、UTYはこれまで考えられていたよりもはるかに広範に、全染色体にわたるゲノム上の約1万箇所に結合していることが明らかとなりました。結合箇所を調べたところ、これらの多くは遺伝子発現制御に重要な領域であるプロモーターおよびエンハンサー領域に位置していました。また、その結合分布はUTXと大きく重なっており、UTYがUTXと機能的に重複しながら、遺伝子発現の制御に関与していることが示されました(図2)。
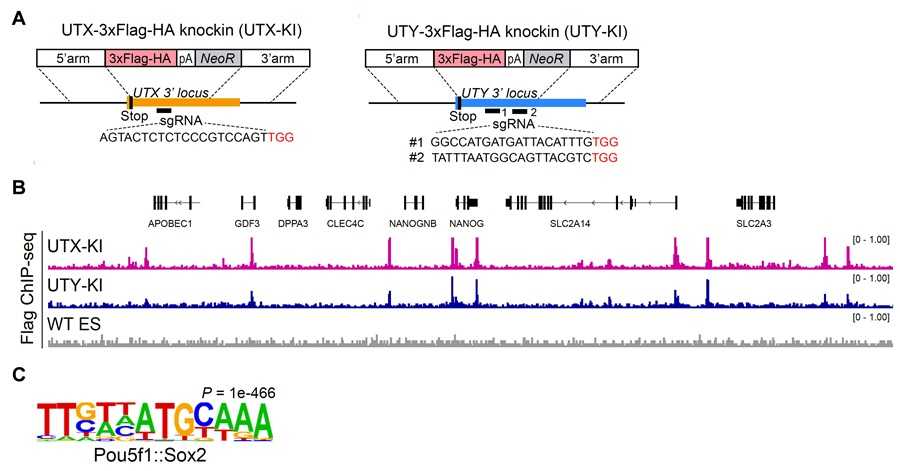
図2 UTXおよびUTYはヒトES細胞において共通の転写制御領域を占有する
UTXおよびUTYのゲノム上の結合を解析するため、CRISPR-Cas9ゲノム編集技術により、各遺伝子座位に3×FLAG-HAタグをノックインしたヒトES細胞株を作製した(A)。Flag抗体によるChIP-seq解析の結果、UTYはUTXと同様に、多能性維持に重要な遺伝子の近くに結合していることが明らかとなった(B)。さらに、UTX/UTYが結合する領域には、多能性維持に重要な転写因子OCT4(Pou5f1)およびSOX2が認識する配列モチーフが多く存在していた(C)。
さらに、UTXとUTYを欠損させた細胞を解析した結果、UTYは酵素として直接作用するのではなく、転写因子やクロマチン制御因子
*8を適切なゲノム領域に配置することで、遺伝子発現を支える役割を担うことが明らかになりました。実際に、UTXとUTYの両方を欠損させると、OCT4やSOX2といった重要な転写因子の結合パターンが広範囲に変化し、それに伴って遺伝子発現に異常が生じました(図3)。
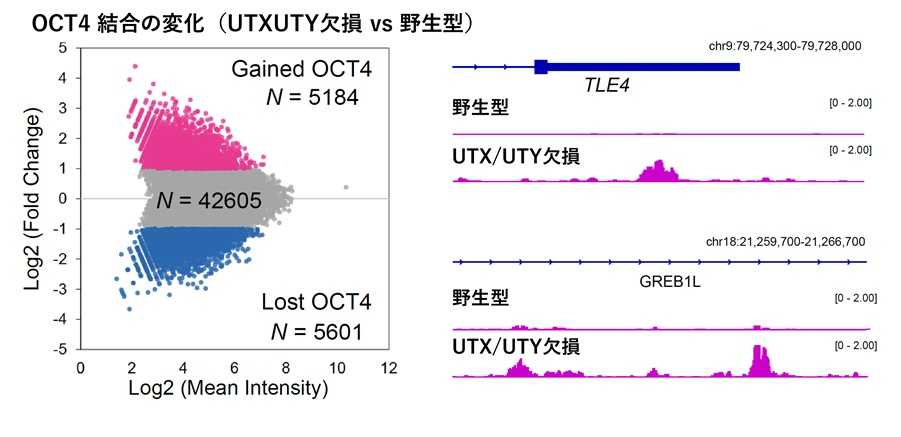
図3 UTX/UTY欠損によるOCT4結合領域の変化
UTXとUTYを同時に欠損させたヒトES細胞では、多能性維持に重要な転写因子OCT4の結合位置が大きく変化した(左)。実際に、本来は結合しない遺伝子領域に新たなOCT4結合が現れることが確認され(右)、UTXとUTYが転写因子OCT4の適切なゲノム局在を維持する役割を持つことが示された。
また、UTXまたはUTYの単独欠損では大きな影響は見られませんでしたが、両方を同時に欠損させると、細胞は未分化な状態を維持できなくなりました。さらに、免疫不全マウス
*9に移植してもさまざまな組織に分化する能力(テラトーマ形成能
*10)を失っており、多能性が著しく損なわれていることが確認されました(図4)。これらの結果から、UTXとUTYは協調して多能性の維持に必須の役割を果たしていることが示されました。
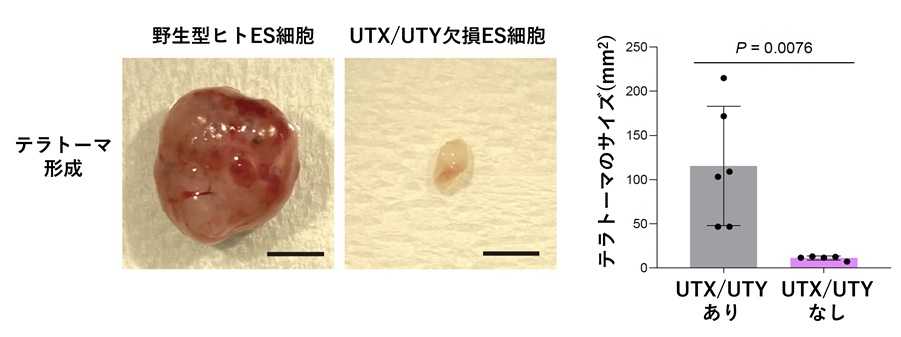
図4 UTX/UTY欠損細胞では多能性が著しく低下する
野生型およびUTX/UTY欠損ヒトES細胞のテラトーマ形成能を評価した。UTX/UTY欠損細胞ではテラトーマ形成能が著しく低下しており、多能性の喪失が示された。(スケールバー、5 mm)
以上の結果は、UTYが酵素活性に依存しない転写制御の中核因子として機能し、ヒト発生を支える新たな分子機構の一端を明らかにしたものです。
今後の展開
本研究は、これまで「小さく、いずれ消える可能性がある」と考えられてきたY染色体が、ヒト発生における遺伝子発現の制御に重要な役割を担うことを示しました。特に、酵素活性をほとんど持たないUTYが転写因子の配置を調整することで遺伝子発現を支えるという発見は、遺伝子制御の新たな理解につながります。さらに本研究は、男性と女性の間で転写制御に違いが存在する可能性を示唆しており、今後は性差に基づく発生制御の分子基盤の解明や、不妊症・発生異常の理解への展開が期待されます。
研究費
本研究は、日本学術振興会 科学研究費助成事業(科研費)(課題番号:JP19K06492、JP22H04699、JP22K06090、JP20H04929、JP20H05395)の支援を受けて実施されました。