顕花植物のゲノムサイズ(核ゲノムDNAの塩基配列長)は、植物種によって大きく異なります。例えば、モデル植物のシロイヌナズナのゲノムサイズは130Mbp(メガ塩基対)、ダイズは1.1Gbp(ギガ塩基対)です。一方で、遺伝子数はシロイヌナズナで約25,000個、ダイズは約60,000個であり、ゲノムサイズほどの違いはありません。つまり、ダイズはシロイヌナズナの10倍の大きさのゲノムを持つものの、遺伝子数は2.5倍程度にとどまります。これは、ダイズゲノムに「遺伝子以外の領域」が多く含まれていることを示しています。この「遺伝子以外の領域」の一つが、トランスポゾン(動くDNA配列)です。トランスポゾンはゲノム内を移動する能力を持ち、その過程でコピー数を増やすことができます。そのため、トランスポゾンは植物のゲノムサイズや構造変化に大きく関わってきたと考えられています。
しかし、トランスポゾンが活発に動くと、重要な遺伝子の中に入り込み、遺伝子の働きを乱す可能性があります。そのため、植物はトランスポゾンの「働きを抑える」仕組みを備えています。実際、ゲノム内の多くのトランスポゾンは、通常はほとんど動かない状態に保たれています。この働きを抑える仕組みには、DNAメチル化やヒストン修飾を介したエピジェネティックな転写抑制が重要な役割を果たしています。
Decreased in DNA methylation 1 (DDM1)は、シロイヌナズナで見出されたクロマチンリモデリング因子です。このDDM1の機能欠損変異体では、ゲノム全体、特にヘテロクロマチンにおけるDNAメチル化が大幅に低下し、トランスポゾンの脱抑制が起こります。それにもかかわらず、シロイヌナズナの場合、変異体は生育可能であり、種子をつけることができます。一方、イネ、トウモロコシ、トマトなどの農作物では、DDM1の機能欠損変異体では、不稔や胚発生致死などの重篤な異常が生じることが知られていました。しかし、DDM1を失ったときの影響が、シロイヌナズナとこれらの作物で、なぜ大きく異なるのかは十分に分かっていませんでした。
Ⅱ.研究の概要・成果
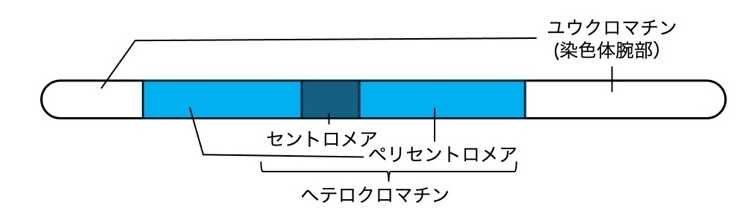
ダイズのゲノムの60%はセントロメアとペリセントロメアからなるヘテロクロマチンで占められており、遺伝子を多く含むユークロマチンは残り40%の染色体腕部に分布しています(図1)。私たちは、このような巨大なペリセントロメアを持つダイズにおいて、DDM1がどのような機能を果たしているのかに興味を持ち、ダイズのDDM1機能欠損変異体を解析しました。
図1ダイズ染色体の模式図
まず、ダイズにはDDM1遺伝子が2つ存在することがわかりました。そこで、ダイズ品種エンレイに変異を誘導した集団から逆遺伝学的にそれぞれの遺伝子の機能欠損変異を見つけました。次に2つのDDM1遺伝子の両方の機能を失った、ダブルミュータントを得るために、どちらか一方のDDM1遺伝子に変異を持つシングルミュータント同士を交配しました。しかし、生育した植物体の中から、ダブルミュータントは得られませんでした。原因を調べたところ、ダブルミュータント由来の種子は見た目にはほぼ正常でしたが、発芽後に成長を停止し、最終的に枯死することが分かりました(図2)。これにより、ダイズにおいてDDM1が発芽後の生育に必須であることが明らかになりました。
図2.ダブルミュータントの発芽後致死性
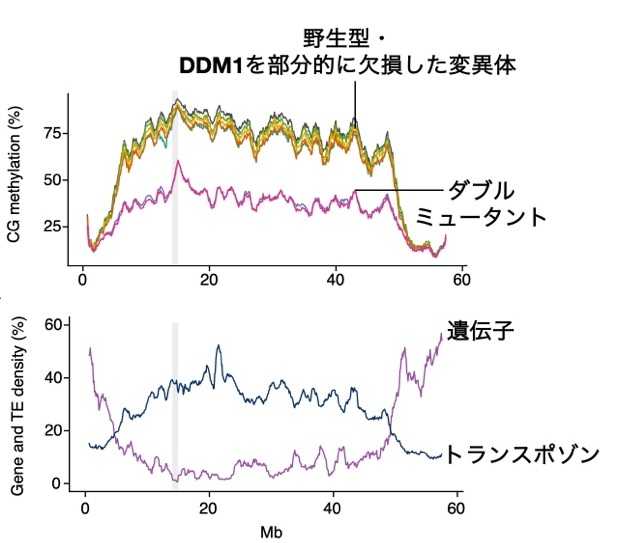
次に、ダブルミュータントにおいてどのような遺伝子の発現が変化しているか調べるため、未熟種子(エダマメの収穫期と同じような時期の種子)を用いて解析を行いました。その結果、約1,950個の遺伝子で発現上昇していることが分かりました。約3,000個のトランスポゾンにおいても発現上昇が見られ、DDM1の機能欠損により大規模なトランスポゾン活性化が起きていることが明らかになりました。さらに、ゲノム全体でDNAのメチル化が大きく低下しており(図3)、ダイズにおいてもDDM1がDNAメチル化の維持に重要であることが確認されました。
図3.ダブルミュータントの大幅なDNAメチル化低下ダイズ第1染色体のデータ。
上:CGサイトのDNAメチル化レベル。
下:トランスポゾンと遺伝子の頻度。
トランスポゾンが多い領域、つまりヘテロクロマチンでのDNAメチル化低下が顕著である。
続いて、DDM1の機能低下によって発現が上昇した遺伝子を詳しく解析しました。その結果、1,400個の遺伝子は、DDM1の機能を部分的に保持している変異体(シングルミュータントなど)でも転写上昇していました。一方、約550個の遺伝子は、DDM1の機能を完全に失ったダブルミュータントでのみ発現が上昇していました。前者の遺伝子は主にユークロマチンに、後者の遺伝子は主にヘテロクロマチンに分布していました。これらの結果から、DDM1量が低下すると、まずユークロマチンに存在する遺伝子の発現異常が生じ、DDM1機能が完全に失われると、ヘテロクロマチンに存在する遺伝子にも影響が広がることが分かりました。
一方、ダブルミュータント特異的に発現が低下した遺伝子の中には、15個の光合成関連遺伝子が含まれていました。また、ダブルミュータントのエダマメでは、デンプン含量の低下も見られました。これらの結果から、DDM1はエダマメ内での光合成関連遺伝子の発現や、種子内の代謝維持に必要であることが示されました。
以上より、ダイズにおいてDDM1が正常な種子発生に不可欠であることが明らかになりました。DDM1の完全な欠損は、大規模なトランスポゾン活性化、DNAメチル化の低下、光合成関連遺伝子の発現低下を引き起こしました。一方で、DDM1機能が部分的に低下した場合には、従来から作用することが知られてきたヘテロクロマチンよりも先に、ユークロマチンに存在する遺伝子の発現異常を引き起こすことが分かりました。ダブルミュータントの致死性が、ダブルミュータント特異的な遺伝子発現変動のみによるのか、ユークロマチン遺伝子の発現変動との相乗効果によるのかは、現段階では分かっていません。本研究では、野生型、DDM1の機能を部分的に保持する変異体、DDM1の機能を完全に失った変異体を比較することで、DDM1がヘテロクロマチンだけでなく、ユークロマチンに存在する遺伝子の発現制御にも関与することを明らかにしました。
DDM1によるユークロマチン遺伝子の発現制御が、ダイズ以外の植物種でも共通して見られる現象なのか、また、その制御が分子レベルでどのように起きているのかは、まだ分かっていません。今後、ゲノムサイズやヘテロクロマチンの割合が異なる植物種でDDM1の機能を比較することで、植物種がそれぞれのゲノム構造に応じて、ヘテロクロマチン制御機構をどのように最適化してきたのかを明らかにできると期待されます。
Ⅳ.研究成果の公表
本研究成果は、2026年5月21日、英国科学誌「The Plant Journal」の電子版に掲載されました。
【論文タイトル】Differential regulation of heterochromatin and euchromatin by GmDDM1 during seed development ensures seedling viability in soybean.
【著者】Ahsen Gers, Kana Shiraishi, Kaoru Tonosaki, Satoru Okamoto, Akito Kaga, Ryota Kuroda, Jun-Ichi Matsuoka, Atsushi Toyoda, Taiji Kawakatsu, Chiho Maruko, Kazuki Takahashi, Keiichi Okazaki, Moeko Okada and Eigo Fukai
【doi】
https://doi.org/10.1111/tpj.70921
Ⅴ.謝辞
本研究はJSPS科研費JP16H06279 (PAGS), JP19K05967, JP23K05163, JP25K01994の助成を受けて行われました。
【用語解説】
(注1)ユークロマチンとヘテロクロマチン
DNAとヒストンなどのタンパク質が形成する複合体をクロマチンといいます。核ゲノムにおいて、クロマチンが密に凝集し高次構造をとっている領域をヘテロクロマチン、クロマチンの凝集がゆるい領域をユークロマチンといいます。前者ではDNAからRNAへの転写が抑制される一方、後者では転写が盛んに行われる傾向があります。
(注2)トランスポゾン(動くDNA配列)
ゲノム内の位置を移動(転移)できるDNA配列の総称。転移先の塩基配列を変えてしまう生物内在の変異源なので、トランスポゾンはヘテロクロマチンとして認識され、転写が抑制されていることが多い一方、過去のトランスポゾンの転移が生物の進化に貢献しているケースもあり、生物の役に立つこともあります。
(注3)エピジェネティクス
DNA配列以外の遺伝情報の総称。例えば、真核生物の核ゲノムDNAのシトシンにはメチル基が付加されることがあり、これをDNAメチル化といいます。同一のDNA配列でも、DNAメチル化されている場合とされていない場合で、DNAからRNAへの転写効率に違いがあることがあります。ヒストンはDNAが巻き付いてクロマチンを構成するタンパク質ですが、ヒストンの種類や修飾によっても転写活性に違いがあることが知られています。
(注4)Decreased in DNA methylation 1 (DDM1)
植物のヘテロクロマチン維持に必要なクロマチンリモデリング因子。植物では広く保存されており、この機能欠損変異体ではゲノム全体、特にヘテロクロマチンのDNAメチル化レベルが低下し、トランスポゾンが活性化されます。機能欠損変異体は、モデル植物のシロイヌナズナ(Arabidopsis thaliana)は正常に生育できる一方、本研究のダイズを含む作物では致死や不稔を示すことが知られています。