【研究背景と内容】
植物は、葉、茎、根の3つの主要器官から成り立っています。その中で、茎は葉や花を支えるだけでなく、根から吸収した水やミネラルを各組織へ運搬する機能や、葉で行われる光合成によって生産された糖を輸送する役割を担っています。また、茎は環境適応や生存競争においても重要な役割を果たしています。例えば、東南アジアなどの季節性の洪水が発生する地域において、茎を伸長させることができる植物は水没から逃れることができます。また、植物が繁茂する地帯では、茎を伸長させることで、光合成に必要な光を多く得ることができるなど、茎の長さは植物の環境適応に深く関わっています。一方、作物の茎の長さを人為的に制御することは重要な育種目標でもあります。例えば風雨による作物の倒伏は収穫量を大きく減少させるため、イネやコムギなどでは茎を短くする育種が行われており、果樹においては収穫効率の向上を目的とした低木化の育種例もあります。このように、植物の茎の伸長制御メカニズムは、生物学的にも興味深い現象であるばかりか、農業的にも重要な現象です。
そこで研究チームは、植物の茎の中でも、特にイネ科作物の茎の伸長メカニズムの解明を目指して、イネを用いた研究を進めました。
イネ、コムギ、オオムギ、トウモロコシをはじめとするイネ科作物の茎は節と節間から構成されており、節間の長さが草丈に影響します(図1)。植物ホルモンの1つであるジベレリン
注1)が、節間の細胞分裂とその後の細胞伸長を促進することで、節間を伸長させることが知られていましたが、分子レベルでの詳細な仕組みについては明らかになっていませんでした。また、節間伸長の開始時期を制御する因子の存在は約50年前に日本人の研究者らによって提唱されていましたが、その実態についても未解明のままでした。
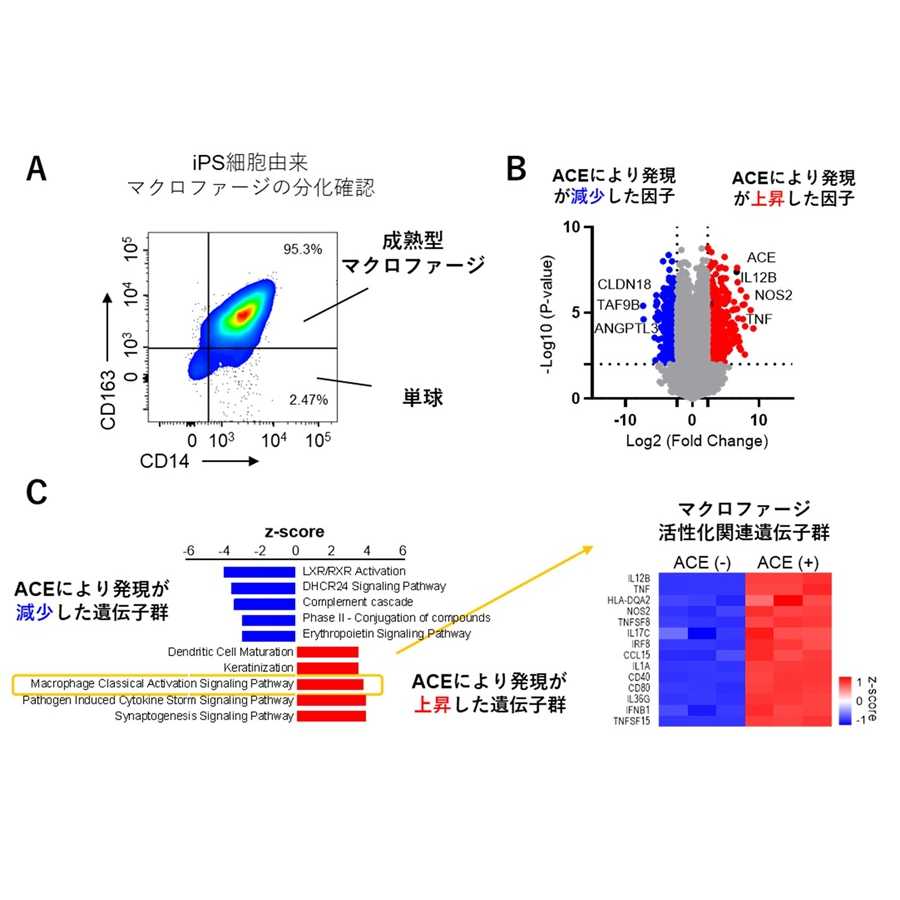
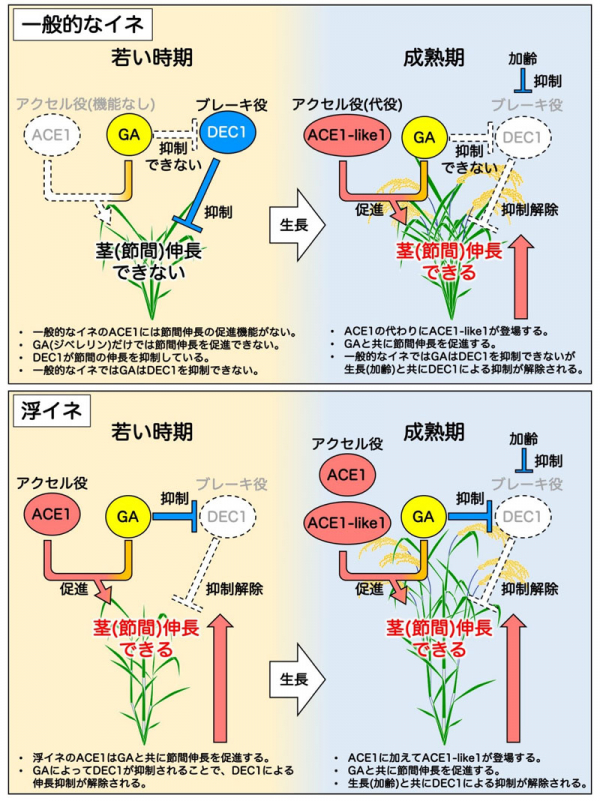
私たちは最初に、一般的な水田で栽培されるイネ(一般的なイネ)と浮イネ
注2)を用いて、ジベレリンと節間伸長の開始時期の関係を調べました。その結果、驚いたことに浮イネは若い時期(栄養生長期の早い段階)にジベレリンを投与すると直ぐに応答して節間伸長を行いましたが、一般的なイネは、若い時期にジベレリンを投与しても節間伸長しませんでした。しかし、一般的なイネも成熟期頃(栄養生長期の後半から生殖生長期)になるとジベレリンに応答した節間伸長をしました。この結果は、ジベレリンだけでは節間伸長の開始に十分ではなく、ジベレリンの応答性を高める因子が節間には存在しており、この因子こそが、長い間、謎であった節間伸長の開始時期を決定する因子であることを示しています。そこで、私たちは遺伝学的手法を用いて、イネの節間においてジベレリンの応答性を制御する因子を探索した結果、2つの因子の存在が明らかとなりました。その後、分子生物学的手法による解析を進めた結果、節間伸長を促進する遺伝子
ACCELERATOR OF INTERNODE ELONGATION1 (
ACE1)と、抑制する遺伝子
DECELERATOR OF INTERNODE ELONGATION1 (
DEC1)を発見しました。
イネ科の茎(節間)伸長では、節間に存在する介在分裂組織と呼ばれる分裂組織が活性化される必要があります(図1)。
ACE1はこの介在分裂組織における細胞分裂を加速させる一方、
DEC1は細胞分裂を抑制する機能があります。浮イネでは若い時期からジベレリン量の増加とACE1タンパク質量の増加が合わさることで節間における細胞分裂が促進されて節間が伸長しますが、一般的なイネでは
ACE1遺伝子に突然変異が入っており、正常なACE1タンパク質が作られないために若い時期にいくらジベレリンを加えても節間伸長をしないことが判明しました(図2)。しかし、一般的なイネにおいても成熟期になると
ACE1遺伝子によく似た遺伝子である
ACE1-like1遺伝子の発現量が増加することで、植物内で合成されたジベレリンに対する応答性が上昇し、それに伴って介在分裂組織における細胞分裂が活性化されることにより節間伸長することが明らかとなりました(図2)。また、一般的なイネのDEC1タンパク質も浮イネのDEC1タンパク質も節間における細胞分裂を抑制する機能がありますが、浮イネではジベレリン量の増加によって
DEC1遺伝子の発現が減少することで節間における細胞分裂が促進されました(図2)。一方、一般的なイネでは、浮イネで観察されたようなジベレリンによる
DEC1遺伝子の発現抑制は起こりませんでした。しかし、一般的なイネにおいても葉齢(主茎の葉数)の増加に伴って
DEC1遺伝子の発現量が徐々に減少していき、成熟期にはその発現が顕著に減少することから、一般的なイネでは成熟期にDEC1による細胞分裂の抑制が解除されることで節間伸長が起きると考えられました。浮イネでは、浮イネ型の
ACE1遺伝子(機能型)と
DEC1遺伝子(若い時期から発現が低下する型)の両方が存在することで、単独で存在する時よりもさらに節間伸長が誘導されたことから、ACE1とDEC1はそれぞれが独立して細胞分裂を制御していることが示されました。また、コムギの近縁種であるミナトカモジグサや、その他にもオオムギやサトウキビにイネの
ACE1遺伝子を導入した植物は節間伸長を促進し、一方でイネの
DEC1遺伝子を発現させたオオムギでは節間の伸長が抑制されたことから、イネにおいて発見した
ACE1遺伝子と
DEC1遺伝子による節間伸長の開始メカニズムはイネ科植物において共通した制御メカニズムであると言うことができ、
ACE1遺伝子と
DEC1遺伝子は今後の作物育種において広く応用できるものと考えられます。
さらに、私たちは、
ACE1遺伝子と
DEC1遺伝子を手掛かりにして、栽培イネとその祖先種である野生イネ
注3)の遺伝子情報を比較することにより、イネにおいてこれらの遺伝子がどのように伝わってきたのかについても解析を行いました。その結果、浮イネが持つ伸長促進効果がある
ACE1遺伝子と一般的なイネが持つ機能が壊れた
ACE1遺伝子、および発現制御が異なる浮イネの
DEC1遺伝子と一般的なイネの
DEC1遺伝子は、野生イネの時点から存在しており、それぞれの遺伝子タイプが、草丈を小さくするための栽培化の過程、または節間伸長による洪水環境適応の過程において選抜されてきたことが明らかになりました。また、浮イネの
ACE1遺伝子と
DEC1遺伝子に加え、これまでに私たちの研究によって明らかにしてきた浮イネの節間伸長に関わる遺伝子
注4)を一般的なイネに導入したところ、洪水環境においても節間を伸長させることで生き残ることができ、最終的には種子を収穫するまでに至りました。これらの結果は、これまでに発見した浮イネの節間伸長に関わる遺伝子に加え、
ACE1遺伝子と
DEC1遺伝子はイネの洪水環境への適応にとても重要な役割を果たしており、これらの遺伝子を組み合わせることで、節間伸長による洪水耐性を獲得したイネを育種できるものと考えています。
【成果の意義】
本研究において発見した、相反する作用を示す2つの因子によって、茎(節間)の伸長を促進するジベレリンと、イネの節間伸長の開始メカニズムとの間に1つの橋を架けることに成功しました。本研究は、浮イネという特殊なイネにおける節間伸長の開始メカニズムに関わる2つの遺伝子の発見に始まり、その後、一般的なイネにおける節間伸長の開始メカニズムの解明、節間の伸長開始を制御する遺伝子の栽培化または環境適応における選抜過程の解明、さらに洪水耐性イネの育種やイネ科作物への応用の可能性を示しました。このことは、浮イネという特殊なイネや野生イネを含めて、「イネ」という植物が持っている多様性に着目して研究を行うことが、いまだ明らかにされていない生物学的現象のメカニズムを解く鍵となることを示しています。今回明らかになった成果を応用することにより、高収量の浮イネ品種の開発や、様々な環境変化に応じてイネやその他のイネ科作物の草丈を調整する技術の確立が期待されます。
【参考図】
図1
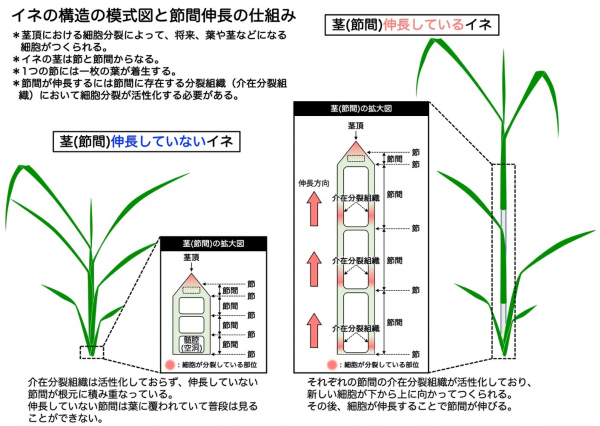
図2
 【用語説明】
【用語説明】
注1):ジベレリン
約100年前に日本人の研究者によって明らかにされた、植物の生長を制御する植物ホルモン(低分子有機化合物)の1つ。植物において細胞分裂や細胞伸長の促進、種子の発芽促進など、多岐にわたる機能を持つ。ジベレリンには複数の分子種が存在しているが、GA1およびGA4が主要な活性型ジベレリンとして機能している。
注2):浮イネ
東南アジアでは、毎年、雨季になると水かさが数メートルにもなる洪水が発生し、この過酷な環境が約4~5ヵ月続く。このような環境では、ほとんどの植物は水没してしまい、呼吸ができず生き残ることができない。一方、バングラデシュやタイなどの東南アジアの国々の洪水が発生する地域では、雨季に向けて「浮イネ」と呼ばれるイネをあらかじめ作付けしている。「浮イネ」は、水の深さに応じて茎(節間)を伸長させるため、洪水環境においても葉先を常に水面より上に出すことで生き残ることができる。そのため、洪水地域に暮らす人々は雨季に洪水が発生してもイネを収穫することができる。品種によってはこの節間の伸長能力が非常に高く、時には草丈が数メートルにおよぶイネも存在する。浮イネの節間伸長には植物ホルモンのうち、エチレンやジベレリンが関わっていることが知られている。
注3):野生イネ
Oryza属のうちアジア栽培イネ(
O. sativa)とアフリカ栽培イネ(
O. glaberrima)を除いた約20種のイネのこと。アジア栽培イネ(
O. sativa)は、野生イネの
O. rufipogonから約8,000年の年月をかけて栽培化されたと考えられている。本研究では野生種として28系統の
O. rufipogonを用いた。
注4):浮イネの節間伸長に関わる遺伝子
過去の研究において植物ホルモンであるエチレンとジベレリンが浮イネの節間伸長を促進することが知られていた。本研究グループではこれまでに、洪水環境によって植物内に蓄積したエチレンに応答して節間伸長を促進する遺伝子
SNORKEL1および
SNORKEL2を発見した(Hattori
et al. 2009,
Nature)。この遺伝子はエチレンによって発現量が上昇し、その結果、節間伸長が引き起こされることが明らかとなった。一般的なイネでは
SNORKEL遺伝子が存在していないためエチレンによる節間伸長が起こらないと考えられた。また、浮イネにエチレンを与えた時に、ジベレリンの生合成量が増加することがこれまでの研究から知られていた。本研究グループは浮イネにおいて、エチレンのシグナルを伝えるタンパク質OsEIL1が、ジベレリン生合成酵素をコードしている遺伝子
GA20ox2の発現量を浮イネにおいて増加させることで、エチレンによるジベレリン量の増加メカニズムを明らかにした(Kuroha
et al. 2018,
Science)。本研究では新たに、ジベレリンに応答して節間伸長の開始を制御する
ACE1遺伝子と
DEC1遺伝子を発見した。
【掲載雑誌、論文名、著者】
雑誌名:Nature
論文タイトル:Antagonistic regulation of the gibberellic acidresponse during stem growth inrice
著者名:Keisuke Nagai, Yoshinao Mori, Shin Ishikawa, Tomoyuki Furuta, Rico Gamuyao, Yoko Niimi, Tokunori Hobo, Moyuri Fukuda, Mikiko Kojima, Yumiko Takebayashi, Atsushi Fukushima, Yasuyo Himuro, Masatomo Kobayashi, Wataru Ackley, Hiroshi Hisano, Kazuhiro Sato, Aya Yoshida, Jianzhong Wu, Hitoshi Sakakibara, Yutaka Sato, Hiroyuki Tsuji, Takashi Akagi and Motoyuki Ashikari
DOI:10.1038/s41586-020-2501-8
論文公開:2020年7月16日0時(日本時間) {2020年7月15日16時(ロンドン時間)}
【著者所属】
名古屋大学生物機能開発利用研究センター(永井啓祐、森欣順、石川慎、Rico Gamuyao、新美陽子、保浦徳典、福田萌莉、芦苅基行)
名古屋大学大学院生命農学研究科(榊原均)
岡山大学農学部(赤木剛士)
岡山大学資源植物科学研究所(古田智敬、久野裕、佐藤和広)
横浜市立大学木原生物学研究所(吉田綾、辻寛之)
情報・システム研究機構国立遺伝学研究所(佐藤豊)
理化学研究所環境資源科学研究センター(小嶋美紀子、竹林裕美子、福島敦史)
理化学研究所バイオリソース研究センター(氷室泰代、小林正智)
農研機構(呉健忠、アキリ亘)
【研究費】
・科学技術振興機構(JST)地球規模課題対応国際科学技術協力プログラム(SATREPS)
「ミャンマーにおけるイネゲノム育種システム強化」(JPMJSA1706)
・科学技術振興機構(JST)戦略的創造研究推進事業チーム型研究(CREST)
「作物の地下茎による栄養繁殖化に向けた基盤技術の開発」(JPMJCR13B1)
・理研-名古屋大科学技術ハブ
・文部科学省新学術領域研究「幹細胞増殖を制御する植物ホルモンの機能解明」(17H06473)
・文部科学省新学術領域研究
「花成ホルモン・フロリゲンを起点とする花形成の「鍵と鍵穴」相互作用の解明」(16H06466)
・科研費 若手研究(B)(16K18565)
・科研費 若手研究(19K15815)
・新農業展開ゲノムプロジェクト(QTL5003)
【謝辞】
浮イネ、野生イネ、オオムギの研究材料は、情報・システム研究機構国立遺伝学研究所およびナショナルバイオリソースプロジェクト(NBRP)から分譲頂きました。